Organelles
Mitochondrium
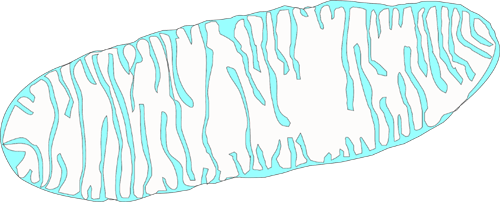
The mitochondrium provides the energy for the cell - Adenosine Trisphosphate (ATP).
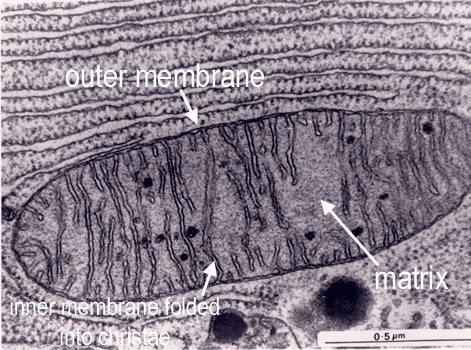
Mitochondria are bounded by two membranes: a smooth outer one,and an inner one which is thrown into folds called cristae.
Endoplasmic reticulum
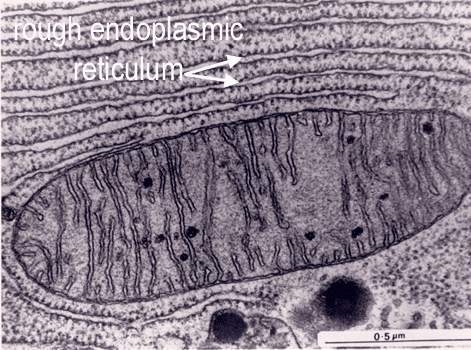
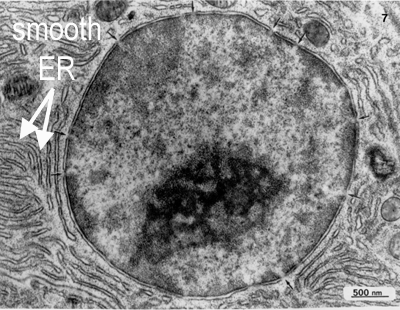
The endoplasmic reticulum is thought to be a single internal membrane that ramifies throughout the cytoplasm enclosing a single internal space (lumen), which may represent more than 10% of the cell volume. The ER occurs in two morphologically distinct forms - smooth (or agranular) endoplasmic reticulum (SER) and rough (or granular) endoplasmic reticulum (RER). The lumen of the RER is continuous with that of the SER.
Golgi-apparatus
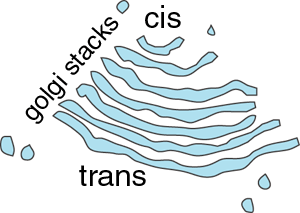
The Golgi apparatus is usually found close to the cell nucleus and consists of one or more stacks of membrane-bound cisternae (sacs). The lumens of these cisternae are separate from each other and from that of the ER.
The Golgi receives synthetic products from the ER, modifies them, and exports them to a variety of destinations. Membrane proteins are collected, sorted and packaged to required destination. For example, secreted proteins are packaged into secretory vesicles and exocytosed. It also produces enzymes that are sorted into lysosomes, to degrade proteins and organelles. It also plays a role in retrieving and recycling proteins.
It has two 'faces' - the cis (receiving or forming or entry) face, and the trans (or maturing or exit) face:
The pathways of endocytosis and exocytosis, are pathways by which proteins are taken up and secreted.
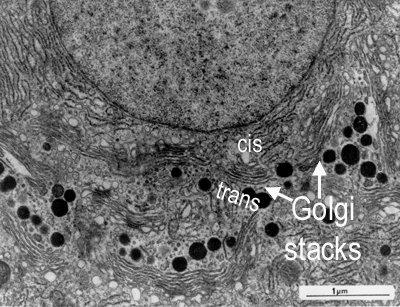
An electron micrograph showing golgi stacks

Secretory vesicles: These are vesicles that can be seen leaving the trans face of the Golgi.
Lysosomes - tiny spherical vesicles (0.2 to 0.4 µm diameter), with an acidic internal PH (ph5.0)-
for degrading proteins. Primary lysosomes are produced by the Golgi apparatus. They form secondary lysosomes by fusing with other membrane-bound
vesicles in the cytoplasm. These vesicles may contain extracellular material that has entered the cell by phagocytosis and require digesting,
or organelles that require degrading (such as in the picture below) because they have reached the end of their active life. They contain
around 40 different types of hydrolytic enzymes, including proteases, nucleases and lipases (they are all acid hydrolases, which need an acid pH to work optimally). This process is also used for degrading internal organelles in a process called 'autophagy' - in which cell organelles are marked for destruction. Lysosomes fuse with the organelles to form secondary lysosomes.
Endosomes - endocytic vesicles formed after endocytosis.
Peroxisomes - derived from rough ER, that contain
enzymes that will form hydrogen peroxide - which phagocytic cells
use to kill bacteria.


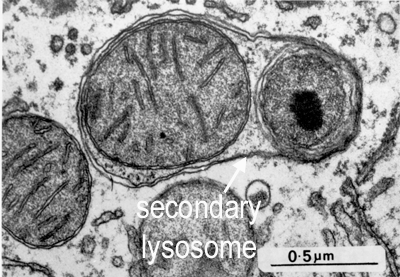
 What organelle is being digested by the secondary lysosome shown in this picture?
What organelle is being digested by the secondary lysosome shown in this picture?